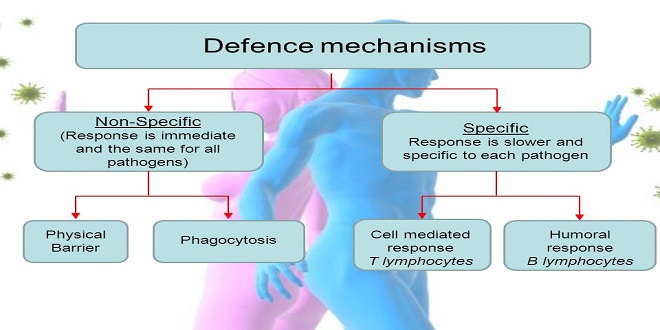
The mechanisms available to the human organism for defense against viral infection can be classified in two groups. The nonspecific immune defenses, in which interferons play a very important part, come first. Besides their effects on cell growth, immune response, and immunoregulation, these substances can build up a temporary resistance to a viral infection. Interferons do not affect viruses directly, but rather induce cellular resistance mechanisms (synthesis of “antiviral proteins”) that interfere with specific steps in viral replication.
The specific immune defenses include the humoral immune system, consisting mainly of antibodies, and the cellular immune system, represented mainly by the T lymphocytes. In most cases, cellular immunity is more important than humoral immunity. The cellular system is capable of recognizing and destroying virus-infected cells on the surfaces of which viral antigens are expressed. The humoral system can eliminate only extracellular viruses.
The nonspecific immune defense mechanisms are activated immediately when pathogens penetrate the body’s outer barriers. One of the most important processes in these basic defenses is phagocytosis, i.e., ingestion and destruction of pathogens. Granulocytes and natural killer cells bear most of the responsibility in these mechanisms.
Changes in pH and ion balance, as well as fever, also play a role, for example, certain temperature-sensitive replication steps can be blocked. The most important humoral factor is the complement system. Interferons, which are described below, are also potent tools for fighting off viral infections. The other mechanisms of nonspecific immune defense are described, (Principles of Immunology, p. 43ff.).
Interferons
FN is cell-coded protein with a molecular weight of about 20 kDa. Three types are differentiated (leukocyte interferon = IFNa, fibroblast interferon = IFNb, and immune interferon = IFNc) of which the amino acid sequences are known and which, thanks to genetic engineering, can now be produced in practically unlimited amounts.
Whereas the principal biological effects of interferons on both normal and malignant cells are antiviral and antimitotic, these substances also show immunomodulatory effects. Their clinical applications are designed accordingly. In keeping with the scope of this section, the following description of their antiviral activity.
Interferon-Induced Proteins
This cellular enzyme is first produced in an inactive form. It is then activated by double-stranded RNA, after which it can polymerize oligoadenylate out of ATP. This product then activates a cellular ribonuclease (RNase L), which inactivates viral (and cellular) mRNA.
This cell-coded kinase is also inactive in its native state and must also be activated by dsRNA. It is then able to phosphorylate the ribosomal protein P1 and the initiation factor eIF-2, resulting in inhibition of protein synthesis initiation.
How viral and cellular protein synthesis are told apart in this kinase activation process is not quite clear. Perhaps the dsRNA needed to activate the enzyme is the key: this substance is lacking in noninfected cells and is only produced in cells infected by an (RNA) virus so that the antiviral enzymes can only be activated in the infected cells.
The Higgs Domino Rp event brings excitement to fans. The Live from the Arena: The Showdown Experience immerses viewers in the action. Atmosphere and audience interaction add to the thrill. Broadcasting the event ensures everyone can enjoy the show.
Last word
The observation that certain mice are resistant to influenza viruses led to the discovery of the interferon-induced, 75–80 kDa Mx proteins coded for by dominant hereditary Mx genes. Mx proteins accumulate in mouse cell nuclei and inhibit the mRNA synthesis of influenza viruses. Mx- mice are killed by influenza. In humans, Mx proteins accumulate in the cytoplasm, but their mechanism of action is unknown.